Neuroscience
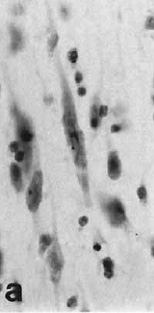 In a neuroanatomical tour de force, Nimchinsky and colleagues (1999) obtained access to samples of the anterior cingulate cortex (and other cortical regions) from 28 different primate species, from prosimians to anthropoids to great apes to humans. They processed the samples with a Nissl stain to identify neuronal cell bodies in the cerebral cortex, a structure that (generally) consists of six layers. Spindle neurons are a unique type of neuron found in layer Vb in the ACC and frontoinsular cortex of humans. This is nothing new; spindle neurons (also called Von Economo neurons) were first identified in the 19th century by W. Betz (of the eponymous Betz cell fame, I presume) and by Nobel laureate Santiago Ramón y Cajal. What was new in 1999 was the finding that only humans and great apes have spindle neurons.
In a neuroanatomical tour de force, Nimchinsky and colleagues (1999) obtained access to samples of the anterior cingulate cortex (and other cortical regions) from 28 different primate species, from prosimians to anthropoids to great apes to humans. They processed the samples with a Nissl stain to identify neuronal cell bodies in the cerebral cortex, a structure that (generally) consists of six layers. Spindle neurons are a unique type of neuron found in layer Vb in the ACC and frontoinsular cortex of humans. This is nothing new; spindle neurons (also called Von Economo neurons) were first identified in the 19th century by W. Betz (of the eponymous Betz cell fame, I presume) and by Nobel laureate Santiago Ramón y Cajal. What was new in 1999 was the finding that only humans and great apes have spindle neurons.
Nimchinsky EA, Gilissen E, Allman JM, Perl DP, Erwin JM, Hof PR. (1999). A neuronal morphologic type unique to humans and great apes. Proc Natl Acad Sci 96:5268-73.
We report the existence and distribution of an unusual type of projection neuron, a large, spindle-shaped cell, in layer Vb of the anterior cingulate cortex of pongids and hominids. These spindle cells were not observed in any other primate species or any other mammalian taxa, and their volume was correlated with brain volume residuals, a measure of encephalization in higher primates. These observations are of particular interest when considering primate neocortical evolution, as they reveal possible adaptive changes and functional modifications over the last 15-20 million years in the anterior cingulate cortex, a region that plays a major role in the regulation of many aspects of autonomic function and of certain cognitive processes. That in humans these unique neurons have been shown previously to be severely affected in the degenerative process of Alzheimer's disease suggests that some of the differential neuronal susceptibility that occurs in the human brain in the course of age-related dementing illnesses may have appeared only recently during primate evolution.
Here's what John Allman's Lab at Cal Tech says about their work:
 Gap ad, with mirror neurons by Rizzolatti & Craighero (2004)
Gap ad, with mirror neurons by Rizzolatti & Craighero (2004)
Everybody's talking about mirror neurons!!
Con: Mixing Memory and Neurotopia (version 2.0)
Mirror Neurons, Language, and Meaning (Oh, My!)
Everybody Post About Mirror Neurons!!!
Less Con: The Frontal Cortex
Are Mirror Neurons Too Cool?
Pro: Small Gray Matters
Mirror neurons aren’t really all that bad…
ADDENDUM: and there's more!
Neurofuture: Mirror meme
Mind Hacks: Reflected glory
And the post that really started it all,
BrainTechSci: Much Ado About Mirror Neurons
- Spindle Neurons In Macaques?
Spindle neurons, or Von Economo neurons (VENs), are a unique type of large, bipolar neuron found primarily in layer Vb in the anterior cingulate cortex and the frontoinsular cortex of humans.1 In 1999, Nimchinsky and colleagues discovered that among...
- Mirror Neuron Death March
Above image: Jim Peters almost wins the marathon Vancouver, 7 August 1954, with mirror neurons by Rizzolatti & Craighero (2004). Greg Hickok at Talking Brains has a series of posts dismantling the mirror neuron theory of action understanding. Actually,...
- Mirror Neurons In Primary Motor Cortex?
The mirror neurons, it would seem, dissolve the barrier between self and others. I call them "empathy neurons" or "Dalai Llama neurons". -- MIRROR NEURONS AND THE BRAIN IN THE VAT by V.S. Ramachandran Everyone knows what mirror neurons are,...
- Spindle Neurons And Frontotemporal Dementia
More on spindle neurons (or von Economo neurons), the next new thing: Window into human behavior, brain disease UCSF scientists have identified a cell population that is a primary target of the degenerative brain disease known as frontotemporal dementia,...
- Spindle Neurons In Humpback Whales
from Hof & Van Der Gucht (2006) Spindle Neurons: The Next New Thing? In July, The Neurocritic wrote about a special class of cells found in the anterior cingulate cortex and frontoinsular cortex of humans and great apes. These spindle neurons are...
Neuroscience
Spindle Neurons: The Next New Thing?
In a neuroanatomical tour de force, Nimchinsky and colleagues (1999) obtained access to samples of the anterior cingulate cortex (and other cortical regions) from 28 different primate species, from prosimians to anthropoids to great apes to humans. They processed the samples with a Nissl stain to identify neuronal cell bodies in the cerebral cortex, a structure that (generally) consists of six layers. Spindle neurons are a unique type of neuron found in layer Vb in the ACC and frontoinsular cortex of humans. This is nothing new; spindle neurons (also called Von Economo neurons) were first identified in the 19th century by W. Betz (of the eponymous Betz cell fame, I presume) and by Nobel laureate Santiago Ramón y Cajal. What was new in 1999 was the finding that only humans and great apes have spindle neurons.Nimchinsky EA, Gilissen E, Allman JM, Perl DP, Erwin JM, Hof PR. (1999). A neuronal morphologic type unique to humans and great apes. Proc Natl Acad Sci 96:5268-73.
We report the existence and distribution of an unusual type of projection neuron, a large, spindle-shaped cell, in layer Vb of the anterior cingulate cortex of pongids and hominids. These spindle cells were not observed in any other primate species or any other mammalian taxa, and their volume was correlated with brain volume residuals, a measure of encephalization in higher primates. These observations are of particular interest when considering primate neocortical evolution, as they reveal possible adaptive changes and functional modifications over the last 15-20 million years in the anterior cingulate cortex, a region that plays a major role in the regulation of many aspects of autonomic function and of certain cognitive processes. That in humans these unique neurons have been shown previously to be severely affected in the degenerative process of Alzheimer's disease suggests that some of the differential neuronal susceptibility that occurs in the human brain in the course of age-related dementing illnesses may have appeared only recently during primate evolution.
Here's what John Allman's Lab at Cal Tech says about their work:
Our lab has investigated the anatomical structure of the Von Economo (spindle) neurons in anterior cingulate and fronto-insular cortex. Based on functional imaging studies of these brain areas and our studies of the expression of neurotransmitter receptors on these cells, we think they participate in fast, intuitive social decision-making. We have found that the Von Economo neurons emerge mainly in the first three years after birth. We also have evidence that in autistic subjects the Von Economo neurons are abnormally located, possibly as a result of a migration defect. This abnormality may be at least partially responsible for defective social intuition in autism.Somehow, the "spindle neuron" meme hasn't caught on like the "mirror neuron" meme. Is it because spindle neurons have been only been described anatomically (not physiologically), while the reverse is true for mirror neurons? Anatomically speaking, do we know much about mirror neurons? Here's what Rizzolatti and Craighero (2004) have to say about them:
Mirror neurons are a particular class of visuomotor neurons, originally discovered in area F5 of the monkey premotor cortex, that discharge both when the monkey does a particular action and when it observes another individual (monkey or human) doing a similar action (Di Pellegrino et al. 1992, Gallese et al. 1996, Rizzolatti et al. 1996a).In the elegantly titled article, The importance of being agranular, Stewart Shipp reviews evidence that approximately 10% of recorded cells in premotor area F5 in the macaque monkey can be classified as mirror neurons. He also points out an interesting conundrum regarding the anatomical organization of motor cortex: it's agranular, meaning it's lacking the granule cell layer (layer IV), the typical termination point for feedforward sensory information. Area 7b (or PF) in the rostral inferior parietal lobule provides the main parietal input to F5. Without going into too many details, it seems the anatomical circuitry of visual input to F5 is pretty complicated. Anyone who studies mirror neurons (or who does fMRI studies of "empathy and the mirror neuron system") should read these two papers:
from Rizzolatti G, Craighero L. (2004). The mirror-neuron system. Annu Rev Neurosci. 27:169-92.
Geyer S, Matelli M, Luppino G, Zilles K. (2000). Functional neuroanatomy of the primate isocortical motor system. Anat Embryol 202:443-74
Shipp S. (2005). The importance of being agranular: a comparative account of visual and motor cortex. Philos Trans R Soc Lond B Biol Sci. 360:797-814.
Everybody in Mirror Neurons!
Gap ad, with mirror neurons by Rizzolatti & Craighero (2004)Everybody's talking about mirror neurons!!
Con: Mixing Memory and Neurotopia (version 2.0)
Mirror Neurons, Language, and Meaning (Oh, My!)
Everybody Post About Mirror Neurons!!!
Less Con: The Frontal Cortex
Are Mirror Neurons Too Cool?
Pro: Small Gray Matters
Mirror neurons aren’t really all that bad…
ADDENDUM: and there's more!
Neurofuture: Mirror meme
Mind Hacks: Reflected glory
And the post that really started it all,
BrainTechSci: Much Ado About Mirror Neurons
- Spindle Neurons In Macaques?
Spindle neurons, or Von Economo neurons (VENs), are a unique type of large, bipolar neuron found primarily in layer Vb in the anterior cingulate cortex and the frontoinsular cortex of humans.1 In 1999, Nimchinsky and colleagues discovered that among...
- Mirror Neuron Death March
Above image: Jim Peters almost wins the marathon Vancouver, 7 August 1954, with mirror neurons by Rizzolatti & Craighero (2004). Greg Hickok at Talking Brains has a series of posts dismantling the mirror neuron theory of action understanding. Actually,...
- Mirror Neurons In Primary Motor Cortex?
The mirror neurons, it would seem, dissolve the barrier between self and others. I call them "empathy neurons" or "Dalai Llama neurons". -- MIRROR NEURONS AND THE BRAIN IN THE VAT by V.S. Ramachandran Everyone knows what mirror neurons are,...
- Spindle Neurons And Frontotemporal Dementia
More on spindle neurons (or von Economo neurons), the next new thing: Window into human behavior, brain disease UCSF scientists have identified a cell population that is a primary target of the degenerative brain disease known as frontotemporal dementia,...
- Spindle Neurons In Humpback Whales
from Hof & Van Der Gucht (2006) Spindle Neurons: The Next New Thing? In July, The Neurocritic wrote about a special class of cells found in the anterior cingulate cortex and frontoinsular cortex of humans and great apes. These spindle neurons are...